UNRAVELING SALMONELLA TYPHIMURIUM: HOW IT MANIPULATES HOST PROTEINS FOR INFECTION SUCCESS

Salmonella enterica subsp. Typhimurium is an enteric, facultative, gram-negative pathogen capable of infecting humans and animals leading to gastroenteritis and systemic infections. It enters via contaminated food and waste and survives the acidic pH of the stomach by activating acid tolerance response (ATR) which provides an inducible pH-homeostatic function and maintains the intracellular pH higher than the extracellular pH to prevent severe acid stock. It adheres to intestinal epithelial cells after passing through the intestinal mucus layer. The adhesion is followed by engaging host signaling pathways leading to major cytoskeletal rearrangements. These disrupt normal epithelial brush border and induce the formation of membrane ruffles that engulf the adherent bacteria in large Salmonella-containing vacuoles (SCVs) where they survive and replicate.
 |
| Model of pathogenesis of Salmonella enterica subsp. Typhimurium |
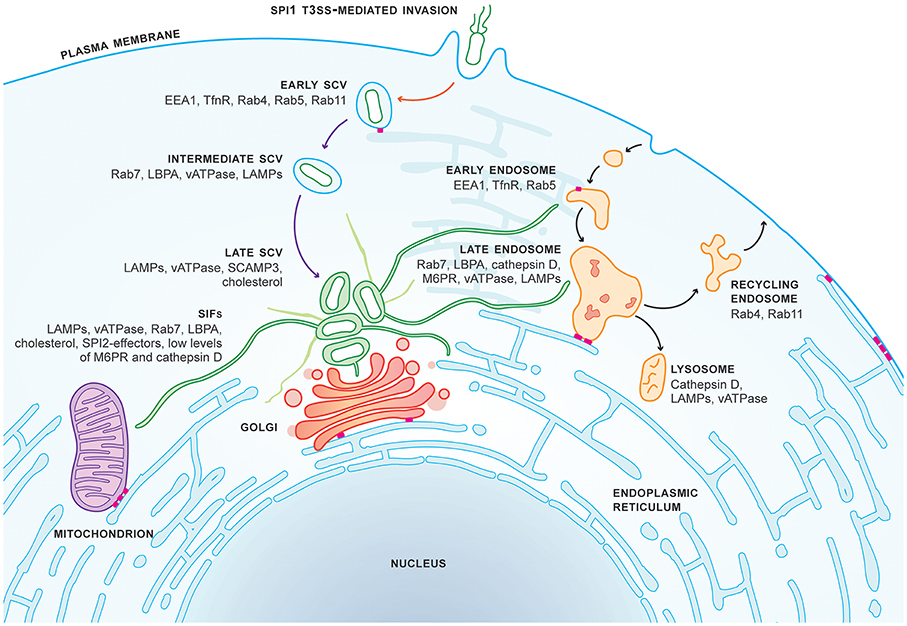
Simultaneously the transmigration of phagocytes to the intestinal lumen was induced by the intestinal epithelium by secretion of tumor necrosis factor-alpha (TNF-α) and interleukin-8 (IL-8). SCVs are prevented from the endocytic pathway by Salmonella-directed changes in host endocytic trafficking. Salmonella induces de novo F-actin meshwork formation which strengthens the integrity of the vacuole. These vacuoles then migrate close to the Golgi apparatus to intercept transport vesicles to obtain nutrients and membrane fragments. The intracellular Salmonella also forms long filamentous membrane structures called Salmonella-induced filaments (SIFs) which can help in increasing the availability of nutrients. All these processes are mediated by the release of toxic effector proteins into the cytoplasm of the infected cell by a two-needle-like type-3-secretion system.
 |
| Maturation of SCV and formation of SIF with various proteins mediating these processes. |
The ability to survive and replicate within the host cell which is important for Salmonella pathogenesis is mediated by the expression of virulence factors located in highly conserved Salmonella pathogenicity islands (SPIs) and virulence plasmid (pSLT). A total of five SPIs were identified: SPI-1 to SPI-5. SPI-1 codes for several proteins that facilitate the invasion of epithelial cells by mediating cytoskeletal rearrangements, SPI-2-coded proteins facilitate the formation of SCV, and the function of the other three SPIs is unknown.
 |
| Genes carried within the five SPIs with their respective functions |
The exact mechanism of how this bacterium modifies the host cellular process still needs to be fully understood. For this scientists employed unbiased systems biology techniques such as unbiased genome-wide genetics and mass spectrometry-based approaches were employed to study host-pathogen protein-protein interactions. One such technique is thermal proteome profiling (TPP) which uses changes in the thermal stability profile of a given protein as an indicator of a change in its biological state. This technique is used to examine how the intracellular bacteria changes the host proteome throughout the infection cycle.
 |
| Workflow of 2D-Thermal proteome profiling |
To distinguish host cell responses from highly immunogenic molecules such as lipopolysaccharides from active bacteria-driven changes to the host proteome, a comparative temporal two-dimensional TPP (2D-TPP) was employed. Murine macrophages (RAW264.7) with live and heat-killed bacteria to ensure both SPI-2 dependent and independent effects on the host proteome were studied. This showed that exposure to live and heat-killed bacteria produced pronounced alteration in host protein abundance in the macrophages but this was largely indistinguishable between the two conditions. This suggests that most protein abundance changes were induced by generic exposure to lipopolysaccharide. However, the changes in protein thermal stability showed less correlation between the two conditions, particularly from the timepoint that coincides with the SPI-2 effector injection and intracellular proliferation (4 hpi) which indicated that the bacterium changes the host cell’s thermal stability landscape. The effect of live bacterium on the host pathways was studied by using relative abundance, and relative thermal stability in a 1D annotation enrichment study. This revealed that the number of host pathways was altered in terms of thermal stability, especially Rab proteins Rab1, Rab5, Rab8, Rab18, and Rab21 which have well-established roles in infection by intracellular bacteria. Thermal destabilization of host ribosomal proteins and thermal stabilization of mitochondrial respiratory chain and SNARE complex proteins were observed. In particular, SNARE proteins are known to regulate SCV dynamics and are targeted by secreted bacterium effectors.
Further thermal stability studies have shown that the abundances of 14 host proteins were altered with proteins like Gbp4 and Ifit3 showing a drastic reduction in the abundance. The Salmonella-induced changes in abundance were found to be pronounced after 4 hpi which correlated with SPI-2 assembly and effector events. Salmonella enterica subsp. Typhimurium injects ubiquitin ligase effectors such as SspH1, SspH2, SlrP, and SopA which are homologous to IpaH9.8 produced by Shigella which ubiquitinates guanylate binding proteins (GBPs) but these did not act on Gbp4 indicating the presence of yet undefined bacterium effector that mediates the reduction of Gbp4 in infected macrophages. It was also identified that 52 proteins exhibited live bacterium-dependent thermal stability changes. It was observed that from 4 hpi, the interferon-induced protein with tetratricopeptide repeats1 (Ifit1) showed drastic thermal destabilization in macrophages. Its stability is regulated by Ifit3 binding which also showed a drastic reduction in abundance at the same timepoint. Acod1 which is mitochondrial cis-aconitate decarboxylase and responsible for itaconate synthesis was also shown to be thermally destabilized. Itaconate is an antimicrobial agent produced by activated immune cells and inhibits host mitochondrial succinate dehydrogenase resulting in succinate accumulation and an enhancement in inflammatory response. Itaconate delivery by Rab32 inhibits bacterium growth in SCV and Acod1 destabilization and thereby reduction in itaconate synthesis may be one of the ways that Salmonella facilitates its intracellular growth and replication.
 |
| Representation of effect of destabilization of Acod1 on itaconate level and its subsequent effects in supporting the bacterial growth |
Using affinity-based mass spectrometry the bacterium effector-host protein-protein interactions were mapped and found that bacterium effector PipB2 regulates Acod1 thermal stability, SifA was found to interact with large ribosomal subunit protein Rpl10a, and thermally destabilizes it and kinase SteC was co-purified with Serbp1 and large ribosomal subunit protein Rpl28a which were both found to be thermally destabilized. In the absence of infection, Serbp1 inhibits ribosomal activity by forming a stable complex with translationally inactive ribosomes. Cellular thermal shift assay (CETSA) revealed that infection of macrophages by Salmonella containing wildtype SteC showed lesser Serbp1 thermal stability than those infected by Salmonella with ΔsteC deletion mutation. Since protein phosphorylation leads to a decrease in thermal stability these data suggest that SteC phosphorylates Serbp1 which was further proved by testing the phosphorylation activity of SteC and inactive SteC in kinase assay with radiolabelled 32P.
In summary, the exploration of Salmonella enterica subsp. Typhimurium reveals its sophisticated mechanisms for manipulating host proteins to establish infection, including the formation of Salmonella-containing vacuoles and the alteration of host cellular processes through the injection of effector proteins. The insights gained from advanced techniques like thermal proteome profiling highlight the dynamic changes in the host proteome during infection, which could pave the way for new therapeutic strategies. Understanding these interactions not only enhances our knowledge of Salmonella pathogenesis but also holds promise for developing targeted interventions against similar intracellular pathogens, ultimately contributing to improved public health outcomes.
REFERENCES:
Fàbrega, A., & Vila, J. (2013). Salmonella enterica serovar Typhimurium skills to succeed in the host: virulence and regulation. Clinical microbiology reviews, 26(2), 308–341. https://doi.org/10.1128/CMR.00066-12
Birk, M. S., Walch, P., Baykara, T., Sefried, S., Amelang, J., Buerova, E., Breuer, I., Vervoots, J., Typas, A., Savitski, M. M., Mateus, A., & Selkrig, J. (2024). Salmonella infection impacts host proteome thermal stability. European journal of cell biology, 103(4), 151448. Advance online publication. https://doi.org/10.1016/j.ejcb.2024.151448
IMAGE SOURCES:
HIPRA, https://static-web.hipra.com/2022-11/salmonella-bacteria-history.png
NCBI, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3623383/bin/zcm9990924170001.jpg
NCBI, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3623383/bin/zcm9990924170002.jpg
ScienceDirect, https://ars.els-cdn.com/content/image/1-s2.0-S0171933524000657-gr1.jpg
ScienceDirect, https://ars.els-cdn.com/content/image/1-s2.0-S0171933524000657-gr3.jpg


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comments
Post a Comment