A JOURNEY INTO MOTION: EXPLORING THE INNER WORKINGS OF BACTERIAL FLAGELLA ROTATION

Mediated by flagella, the motility of bacteria forms a cornerstone in their survival, allowing them to colonize new environments and pursue nutrients. This whip-like appendage plays an important role in motility and chemotaxis and cell adhesion, invasion, biofilm formation, protein export, and virulence of bacteria providing an advantage over non-motile counterparts. It is a complex structure made of more than 20 different proteins. The bacterial chemotaxis depends on the bidirectional flagellar rotation involving the switching the rotation between clockwise and anticlockwise directions allowing the bacteria to travel at a particular path and to change to new trajectories.
Until recently the exact mechanism of this switching of the direction of rotation, torque transfer, and its regulation by response regulators is unknown. Scientists used cryoEM (cryogenic electron microscopy) to reveal the various conformational changes happening in flagella to understand the mechanisms of these processes.
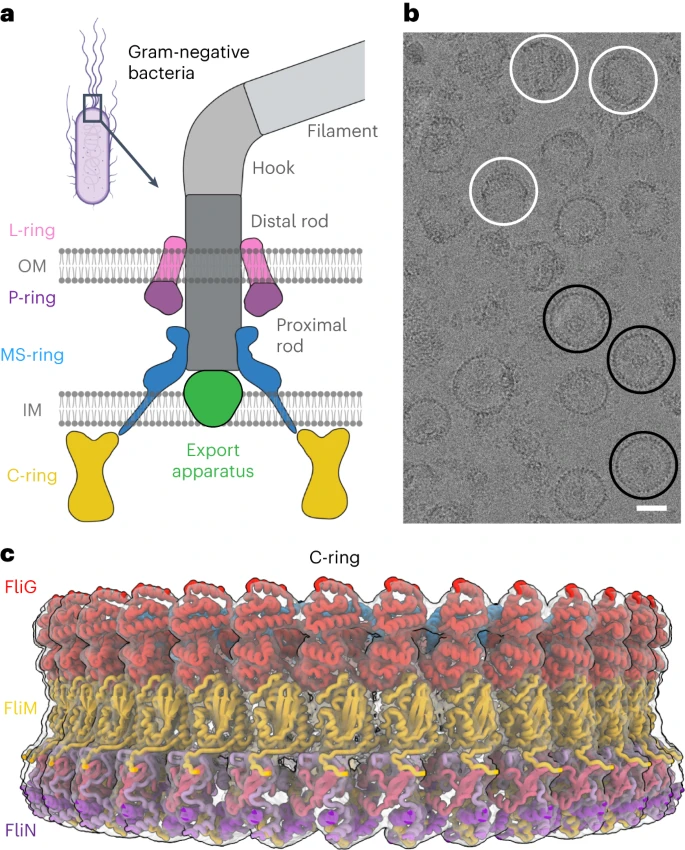
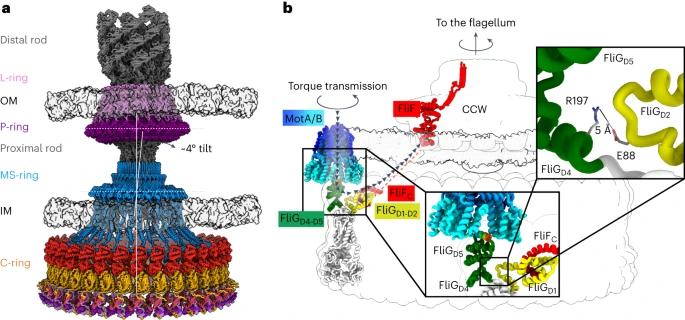
Flagella consists of three parts: filament which is involved in propulsion, and hook which connects the filament to the basal body which usually consists of four rings L-ring, P-ring, MS-ring, and C-ring in case of gram-negative bacteria (L=lipid, P=peptidoglycan, MS=membrane-super membrane, C=cytoplasm) and two rings: peptidoglycan ring and plasma membrane ring in gram-positive bacteria.
 |
| Structure of flagella |
These rings make up the flagella motor which is involved in switching the rotation between clockwise and anticlockwise directions and hence acting as a switch and this is made of multiple copies of protein subunits of FliG, FliM, and FliN. to understand the mechanisms involved in switching, the structure of wild-type C-ring from Salmonella enterica serovar typhimurium was elucidated using cryoEM from the purified 6 MDa complex consisting of both MS-ring and C-ring which along with data from docking Alphafold models revealed that switch subunits are organized into layers made of FliF514–560/FliG1–331 in two layers at the top at the side of C-ring which faces the MS-ring, FliM52–237 (FliMmid) in the middle, and FliM257–330(FliMC)/FliN45/59/63–137 in a 3:1 ratio at the bottom. It also revealed that FliG contains five domains when assembled into switch contrary to previous knowledge of having only three domains.
 |
| CryoEM image of wild-type MS- and C-rings |
 |
| Representation of C-ring |
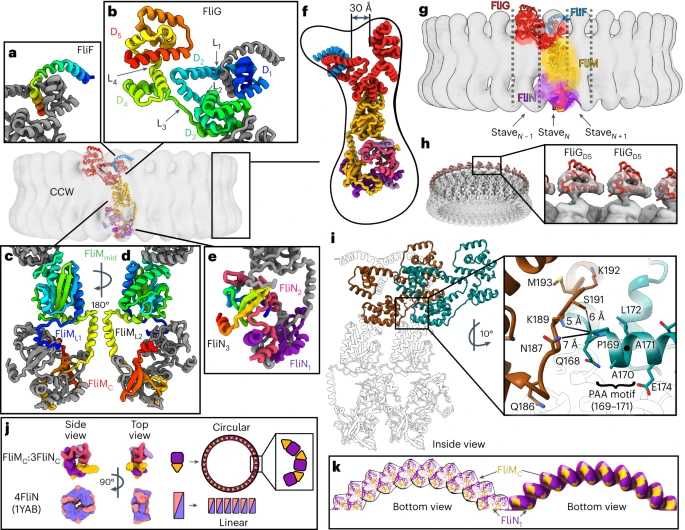
The purified C-ring under physiological conditions rotates only in the anticlockwise direction and was considered to be CCW pose. In this conformation, the individual FliG forms a v-shape with an inner ring of FliFC, FliGD1, and FliGD2 and an outer ring of FliGD5 separated by 30 Å cleft. FliFC forms a curved helix that extends radially from the MS-ring on the membrane side and interacts strongly with FliGD1. FliGD1 extends into FliGD2 to form an armadillo motif, an α-helical hairpin that permits rotations. This motif completes the fold of the next FliGD1 to form an intercalated structure. The armadillo motifs of FliGD3 and FliGD4 intercalate around the ring via domain swaps below these upper rings. FliGD4 also forms the base of the cavity. FliGL3 stabilizes the position of FliGD2 and interacts with a pro-ala-ala (PAA) sequence motif and deletion of this PAA motif results in predominate clockwise rotation of the flagellar motors.
 |
| CCW pose of the switch |
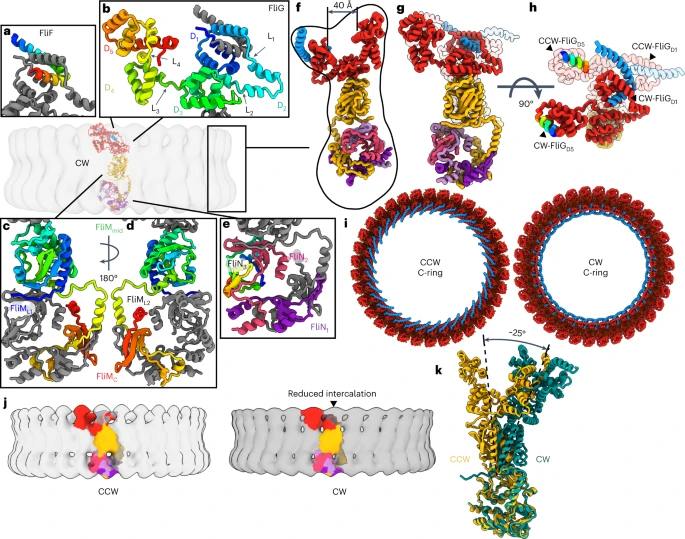
The conformation of the flagellar motor responsible for clockwise rotation, termed as CW pose had major domain rearrangements, particularly in FliG. FliFC–FliGD1 and FliGD2 of the inner ring and FliGD5 of the outer ring are rotated by -180o. Due to this FliGD2 binds to different subunits thereby altering domain swaps. This reverses the orientation of FliGD5 which binds to the MotA/B stator and FliGD1 binding to the MS-ring and this results in an increase in the size of the cleft from 30 Å to 40 Å. These altered domain swaps facilitate C-ring assembly and support directional cooperativity. The loss of the PAA motif shortens the FliGL3 linker, adjusts the position between FliGD3 and FliGD4, moves FliGD2 to eliminate a domain swap, and allows FliM to rotate.
 |
| CW pose of the switch |
 |
| Comparison of the top view of CCW and CW ring showing the reversed orientation of FliFC helix |
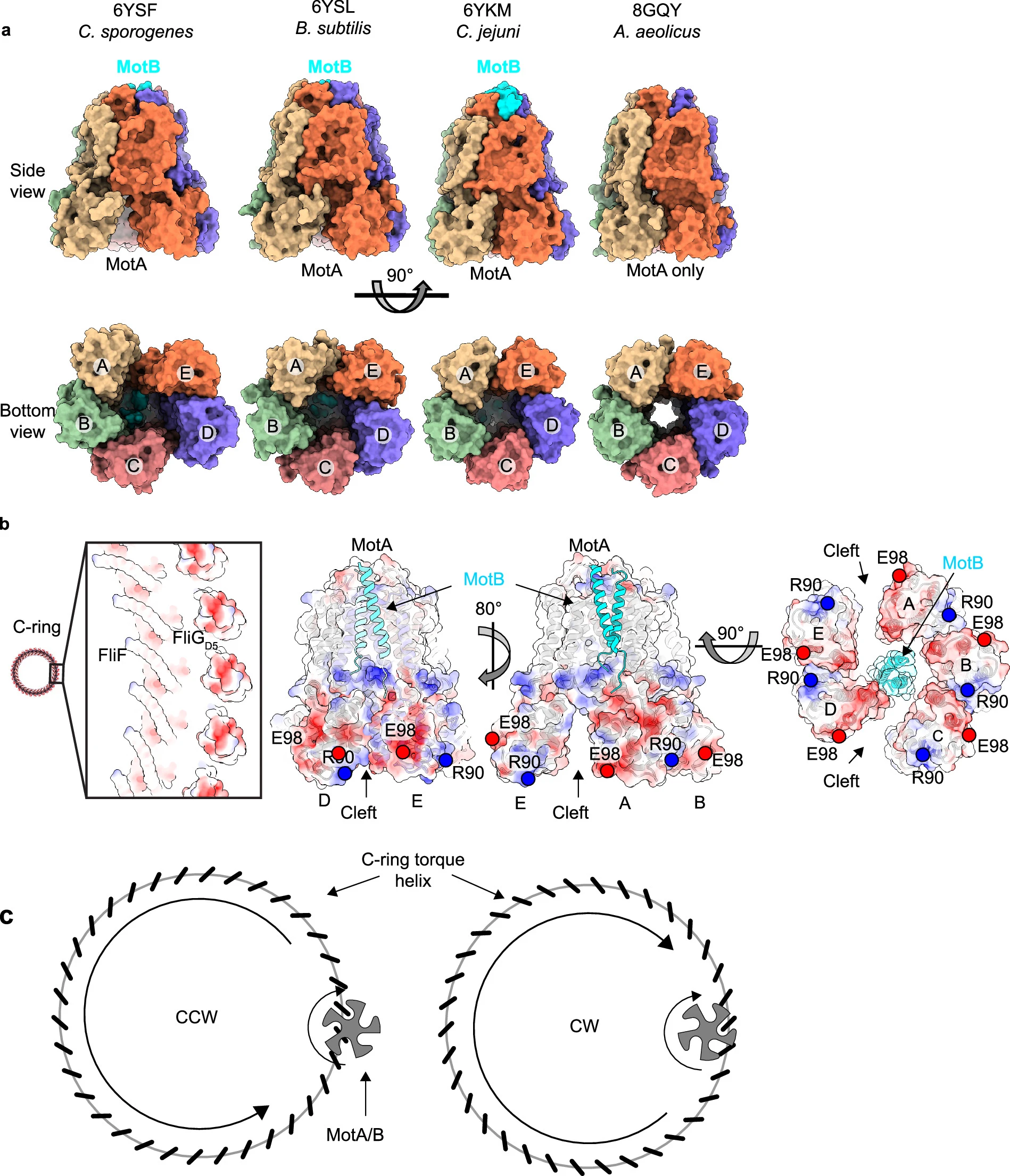
It was found that torque enters from the C-ring at the outer FliGD5. In the CW pose, the rotation of FliGD5 presents the MotA/B binding site to the inside of the ring along with the transmission of torque in the same direction. The C-ring transfers torque to the MS-ring and filament via the MotA/B stator. The FliGD5 interacts with the MotA/B stator and transmits to FliGD1 whose direct interaction with FliF turns the MS-ring and the filament. The cryoEM and Alphafold data revealed that MS-ring and C-ring stack over each other with a tilt of 4o. This tilt results in wobble rotation of the flagella and this symmetry mismatch allows for efficient rotation.
 |
| 1) Model of the flagellar motor of gram-negative bacteria showing the between MS- and C-ring 2) torque transmission pathway transmission from C-ring to MS-ring and filament via MotA/B stator |
 |
| Bidirectional MotA/B dependent movement of C-ring |
In conclusion. cryoEM data revealed that torque produced by the transmembrane electrochemical gradient induces the rotation and gets transmitted from the C-ring to the MS-ring. It suggested the conformational difference between the CW pose and CCW pose on the FliG subunit that moves the binding site for MotA/B from facing inward in the CW pose to facing outward in the CCW pose which changes the direction of rotation of the C-ring. This also explains cooperativity which induces all the subunits to adjust to the same pose. The CW data revealed a binding site in 40 Å cleft for the regulatory proteins involved in changing the direction of rotation. 30 Å cleft in CCW pose may also have a binding site for regulatory proteins.
The study of bacterial flagella rotation reveals a fascinating world of molecular intricacies essential for bacterial survival and adaptation. This enhances our understanding of microbial motility, from fundamental biology to potential applications in biotechnology and medicine. Uncovering flagellar dynamics opens doors to new insights and innovations that could shape future scientific advancements and solutions for various challenges.
REFERENCES:
Singh, P.K., Sharma, P., Afanzar, O. et al. CryoEM structures reveal how the bacterial flagellum rotates and switches direction. Nat Microbiol 9, 1271–1281 (2024). https://doi.org/10.1038/s41564-024-01674-1
IMAGE SOURCES:
Eberly College of Science, https://ecos-appdev-production.s3.amazonaws.com/science_site/s3fs-public/inline-images/2017news-e-coli.jpg


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comments
Post a Comment